
Jarmo Laitinen1, Paavo Ojanen2, Kaisu Aapala3, Juha-Pekka Hotanen4, Aira Kokko3, Pekka Punttila3, Sakari Rehell5, Juha Tiainen2,4 ja Harri Vasander2
1Oulun yliopisto, 2Helsingin yliopisto, 3Suomen ympäristökeskus, 4Luonnonvarakeskus, 5Metsähallitus Luontopalvelut
Julkaistu 24.11.2020
Johdanto
Suot ovat hyvin monimuotoisia elinympäristöjä (ks. Soiden kasvillisuus). Niillä vaihtelevat sekä märkyys että ravinteisuus ja niiden seurauksena myös avoimuus ja puustoisuus. Lisäksi suurilmastollisten erojen vuoksi suot ovat erilaisia Etelä- ja Pohjois-Suomessa. Ojitus hävittää useimmat luonnontilaisten soiden piirteet hyvin tehokkaasti, mutta jopa korostaa joitain piirteitä. Ojitus on myös keskittynyt erityisesti reheville sekä puustoisille soille ja toisaalta Etelä- ja Keski-Suomeen (ks. Soiden käyttö Suomessa ja Suoluontotyyppien uhanalaisuus). Kun vielä erilaisille eliölajiryhmille hyvin erilaiset elinympäristön ominaisuudet ovat tärkeitä, soiden ja niiden ojituksen merkitys eliölajeille ja niiden uhanalaisuudelle riippuu voimakkaasti tarkasteltavasta lajiryhmästä (Aapala 2001b). Ojituksen yleinen vaikutus on, että se samankaltaistaa soita ja siksi hävittää suoluonnolle ominaista monimuotoisuutta.
Soiden eliölajien uhanalaisuus
Uusimman uhanalaisarvion (Hyvärinen ym. 2019) mukaan eliölajeista, joiden ensisijainen elinympäristö on suo, 120 on uhanalaisia eli äärimmäisen uhanalaisia (CR), erittäin uhanalaisia (EN) tai vaarantuneita (VU) (taulukko 1). Erityisen paljon uhanalaisia lajeja soilla on hyönteisissä, putkilokasveissa ja sammalissa. Ensisijaisten suolajien osuus kaikista Suomen uhanalaisista lajeista on vain 4,5 %, mikä johtuu siitä, että suot ovat useimmiten vähälajisia elinympäristöjä. Uhanalaisten lajien osuus kaikista ensisijaisista suolajeista on 11 %, mikä on samaa tasoa kuin uhanalaisten lajien osuus Suomen kaikesta lajistosta keskimäärin. Lisäksi 129 uhanalaiselle lajille suo on toissijainen elinympäristö (niiden pääelinympäristö on jokin muu, mutta ne esiintyvät myös soilla). Näistä lajeista puolet on ensisijaisesti metsälajeja ja lähes kolmannes vesistöjen ja rantojen lajeja. Erityisesti luonnontilaiset korvet ovatkin monille vanhojen metsien lajeille suotuisia elinympäristöjä kostean pienilmaston, monimuotoisen puuston ja lahopuujatkumon ansiosta (Aapala 2001a).
Taulukko 1. Uhanalaisten (CR, EN, VU) ja silmälläpidettävien (NT) ensi- ja toissijaisten suolajien lukumäärä (n) lajiryhmittäin (Hyvärinen ym. 2019). Osuus (%) kertoo, kuinka suuri osa kyseisessä lajiryhmässä arvioiduista suolajeista on uhanalaisia tai silmälläpidettäviä. Tiedot on haettu Punaisen kirjan verkkopalvelusta (https://punainenkirja.laji.fi/) 17.2.2020. Uhanalaisuusluokat: CR = äärimmäisen uhanalaiset, EN = erittäin uhanalaiset, VU = vaarantuneet.
|
|
Ensisijaiset suolajit |
|
Toissijaiset suolajit |
||||||||
|
Lajiryhmä |
Uhanalaisia |
|
Silmälläpidettäviä |
|
Uhanalaisia |
|
Silmälläpidettäviä |
||||
|
|
n |
% |
|
n |
% |
|
n |
% |
|
n |
% |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Hyönteiset |
54 |
8 |
|
68 |
11 |
|
36 |
6 |
|
53 |
9 |
|
Putkilokasvit |
20 |
17 |
|
11 |
9 |
|
19 |
49 |
|
7 |
18 |
|
Sammalet |
19 |
15 |
|
5 |
4 |
|
31 |
31 |
|
14 |
14 |
|
Sienet ja jäkälät |
13 |
13 |
|
7 |
7 |
|
28 |
27 |
|
21 |
20 |
|
Linnut |
8 |
33 |
|
6 |
25 |
|
11 |
20 |
|
9 |
16 |
|
Nilviäiset |
2 |
67 |
|
1 |
33 |
|
0 |
0 |
|
1 |
7 |
|
Hämähäkkieläimet |
4 |
4 |
|
11 |
10 |
|
0 |
0 |
|
2 |
2 |
|
Nisäkkäät |
0 |
0 |
|
0 |
0 |
|
2 |
15 |
|
2 |
15 |
|
Matelijat ja sammakkoeläimet |
0 |
0 |
|
0 |
0 |
|
2 |
20 |
|
0 |
0 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Yhteensä |
120 |
11 |
|
109 |
10 |
|
129 |
13 |
|
109 |
11 |
Lettojen lajeista melkein puolet on uhanalaisia tai silmälläpidettäviä (NT), kun keskimäärin kaikista ensisijaisista suolajeista viidennes on sellaisia (taulukko 2). Lettojen merkitys sekä runsaslajisena että ojituksen myötä erityisen voimakkaasti uhanalaistuneena elinympäristönä näkyy siinä, että uhanalaisista ja silmälläpidettävistä ensisijaisista suolajeista kolmannes on lettolajeja, vaikka Suomen soista vain muutama prosentti on lettoja (Ilvessalo 1957, Korhonen ym. 2017).
Taulukko 2. Uhanalaisten (CR, EN, VU) ja silmälläpidettävien (NT) ensisijaisten suolajien määrä (n) ja osuus (%) arvioiduista lajeista elinympäristöittäin (Hyvärinen ym. 2019). Lisäksi on esitetty uhanalaisten ja silmälläpidettävien ensisijaisten suolajien kokonaismäärän jakaantuminen eri suoelinympäristöihin (Osuus). Lettoihin on yhdistetty lettojen (avosoita) lisäksi lettorämeet ja lettokorvet. Nevoihin on yhdistetty kaikki lettoja karummat avosuot. Rämeisiin on yhdistetty aidot puustoiset rämeet ja sekatyypin rämeet. Korpiin on yhdistetty aidot puustoiset korvet ja sekatyypin korvet. Tiedot on haettu Punaisen kirjan verkkopalvelusta (https://punainenkirja.laji.fi/) 17.2.2020. Uhanalaisuusluokat: CR = äärimmäisen uhanalaiset, EN = erittäin uhanalaiset, VU = vaarantuneet.
|
|
Uhanalaisia |
Silmälläpidettäviä |
Osuus |
||
|
n |
% |
n |
% |
||
|
letot |
55 |
31 |
26 |
15 |
35 % |
|
nevat |
23 |
10 |
16 |
7 |
17 % |
|
rämeet |
14 |
7 |
35 |
17 |
21 % |
|
korvet |
17 |
11 |
11 |
7 |
12 % |
|
tarkemmin määrittelemätön |
11 |
3 |
21 |
6 |
14 % |
|
120 |
11 |
109 |
10 |
100 % |
|
Soilla ensi- ja toissijaisesti esiintyviä valtakunnallisesti uhanalaisia ja silmälläpidettäviä lehti- ja maksasammalia tarkasteltaessa korostuvat elinympäristöinä runsaslajiset letot ja korvet (taulukot 3 ja 4). Korpien lajeista suuri osa on myös vanhojen kangasmetsien lajeja. Rämeiden ja nevojen merkitys uhanalaisten sammallajien elinympäristöinä on pienempi, koska ne ovat melko vähälajisia ja niitä on myös säilynyt melko paljon ojittamattomina (ks. Soiden käyttö Suomessa, taulukko 2). Lisäksi rämeiset piirteet voivat säilyä melko pitkään ja aluksi jopa vahvistua metsäojituksen jälkeen (ks. Soiden kasvillisuus ja Suoluontotyyppien uhanalaisuus).
Taulukko 3. Ensisijaisesti ja toissijaisesti suolla esiintyvät uhanalaiset (CR, EN, VU) ja silmälläpidettävät (NT) lehtisammallajit (Bryophyta) karkeasti suoelinympäristöittäin (Hyvärinen ym. 2019). Korpisarakkeessa merkintä v viittaa lajeihin, jotka esiintyvät korpien tai kivennäismaiden vanhoissa metsissä tai joiden kasvupaikkoihin liittyy vanhan metsän piirteitä. Ojitukseen, turpeennostoon ja pellonraivaukseen liittyvät uhanalaisuuden syyt on esitetty kullekin lajille: O/o = ojitus ja turpeenotto ensisijainen / yksi uhanalaisuuden syy, Pr/pr = pellonraivaus ensisijainen / yksi uhanalaisuuden syy. Merkinnät >——— ja ————< tarkoittavat, että laji esiintyy myös kyseisessä ympäristössä. Merkintä >— nevojen kohdalla tarkoittaa esiintymistä rehevimmillä nevoilla. Tiedot on haettu Punaisen kirjan verkkopalvelusta (https://punainenkirja.laji.fi/) 17.2.2020. Uhanalaisuusluokat: CR = äärimmäisen uhanalaiset, EN = erittäin uhanalaiset, VU = vaarantuneet.
|
LETOT |
NEVAT |
RÄMEET |
KORVET |
|
Ensisijaisesti soilla esiintyvät lajit |
|||
|
Amblyodon dealbatus O pr |
|
|
|
|
Campylium laxifolium O |
|
|
|
|
Hamatocaulis vernicosus O |
|
|
|
|
Ptychostomum longisetum Pr o |
|
|
|
|
Meesia longiseta Pr o |
>—— |
|
|
|
Sphagnum contortum O |
|
|
|
|
Sphagnum subnitens O |
>—— |
|
|
|
|
Sphagnum affine.O |
|
>—————————————— |
|
|
Sphagnum molle O |
|
|
|
|
Pohlia sphagnicola O |
>———— |
|
|
——————————————< |
——————————————< |
————< |
Splachnum melanocaulon O |
|
|
|
|
Herzogiella turfacea v O |
|
|
|
|
Mnium spinosum o |
|
|
|
|
Plagiothecium latebricola v o |
|
|
|
|
Pseudocampylium radicale v O |
|
|
|
|
Timmia megapolitana |
|
Toissijaisesti soilla esiintyvät lajit |
|||
|
Campyliadelphus elodes o |
|
|
|
|
Drepanocladus angustifolius o |
|
|
|
|
Drepanocladus lycopodioides O pr |
|
|
|
|
Drepanocladus sendtneri O |
|
|
|
|
Palustriella falcata o |
|
|
|
|
Pseudoscleropodium purum Pr |
|
|
|
|
Hypnum bambergeri |
|
|
|
|
Orthothecium lapponicum |
|
|
|
|
Dicranella grevilleana o |
|
|
|
|
Cratoneuron filicinum |
|
|
|
|
Philonotis calcarea O |
|
|
|
|
———————————————< |
Hamatocaulis lapponicus O |
|
|
|
———————————————< |
Sphagnum inundatum O |
|
|
|
|
|
|
Polytrichum pallidisetum v |
|
|
|
|
Callicladium haldanianum |
|
|
|
|
Tayloria tenuis O |
|
|
|
|
Mnium lycopodioides o |
|
Suoelinympäristö tarkemmin määrittelemätön: Dicranum laevidens |
|||
Taulukko 4. Ensisijaisesti ja toissijaisesti suolla esiintyvät uhanalaiset (CR, EN, VU) ja silmälläpidettävät (NT) maksasammallajit (Marchantiophyta) karkeasti suoelinympäristöittäin (Hyvärinen ym. 2019). Korpisarakkeessa merkintä v viittaa lajeihin, jotka esiintyvät korpien tai kivennäismaiden vanhoissa metsissä tai joiden vastaaviin kasvupaikkoihin liittyy vanhan metsän piirteitä. Merkintä lä viittaa lajeihin, jotka ovat ensisijaisesti lähteikköjen lajeja. Ojitukseen ja turpeennostoon liittyvät uhanalaisuuden syyt esitetty kullekin lajille: O/o = ojitus ja turpeenotto ensisijainen / yksi uhanalaisuuden syy. Pellonraivausta ei ole arvioitu yhdenkään maksasammallajin uhanalaistumisen syyksi. Merkinnät >——— ja ————< tarkoittavat, että laji esiintyy myös kyseisessä ympäristössä. >— nevojen kohdalla tarkoittaa esiintymistä rehevimmillä nevoilla. Tiedot on haettu Punaisen kirjan verkkopalvelusta (https://punainenkirja.laji.fi/) 17.2.2020. Uhanalaisuusluokat: CR = äärimmäisen uhanalaiset, EN = erittäin uhanalaiset, VU = vaarantuneet.
|
LETOT |
NEVAT |
RÄMEET |
KORVET |
|
Ensisijaisesti soilla esiintyvät lajit |
|||
|
Mesoptychia bantriensis O |
|
|
|
|
Schistochilopsis grandiretis O |
|
|
|
|
Biantheridion undulifolium o |
>—— |
|
|
|
Heterogemma capitata o |
>—— |
|
|
|
Moerckia hibernica sensu lato O |
>—— |
|
>——————————————————— |
|
|
Heterogemma laxa O |
>————— |
|
|
|
————————————< |
—————< |
Odontoschisma denudatum v o |
|
|
|
|
Harpanthus scutatus v o |
|
Toissijaisesti soilla esiintyvät lajit |
|||
|
Riccardia multifida lä O |
|
|
>——————————————————— |
|
|
|
Mylia taylorii O |
|
|
|
|
Calypogeia fissa O |
>—————————————————— |
|
|
|
—————< |
Nowellia curvifolia v o |
|
|
|
|
Trichocolea tomentella lä o |
|
|
|
|
Aneura maxima lä o |
|
|
|
|
Aneura mirabilis lä o |
|
|
|
|
Anastrophyllum michauxii v o |
|
|
|
|
Calypogeia suecica v o |
|
|
|
|
Cephalozia lacinulata v o |
|
|
|
|
Cephalozia macounii v o |
|
|
|
|
Crossocalyx hellerianus v o |
|
|
|
|
Fuscocephaloziopsis catenulata v o |
|
|
|
|
Geocalyx graveolens v O |
|
|
|
|
Liochlaena lanceolata v o |
|
|
|
|
Lophocolea bidentata lä o |
|
|
|
|
Lophozia ascendens v o |
|
|
|
|
Lophozia ciliata v o |
|
|
|
|
Lophozia guttulata v o |
|
|
|
|
Riccardia palmata v o |
|
|
|
|
Scapania apiculata v o |
|
|
|
|
Scapania carinthiaca. v o |
|
|
|
|
Syzygiella autumnalis v o |
|
|
|
|
Tritomaria exsectiformis v o |
|
Suoelinympäristö tarkemmin määrittelemätön: Conocephalum salebrosum o, |
|||
Soilla ensi- ja toissijaisesti esiintyvien uhanalaisten putkilokasvilajien elinympäristöinä korostuvat sammalten tapaan erityisesti letot (taulukko 5). Myös korpien lajeja on runsaasti.
Uhanalaisten ja silmälläpidettävien ensi- ja toissijaisten suolintulajien elinympäristöjen tärkeitä ominaispiirteitä ovat suotyypin sijaan suon avoimuus tai harvapuustoisuus sekä märkyys (taulukko 6). Uhanalaisista ja silmälläpidettävistä suolintulajeista ensisijaiset suolajit esiintyvät pääasiassa Suomen pohjoisosissa, mutta toissijaisista lajeista monet ovat Suomen eteläosissakin esiintyviä avoimien ympäristöjen lajeja (taulukko 6). Puustoiset suot, erityisesti korvet, taas ovat tyypillisesti metsälajeiksi luokiteltujen lintujen elinympäristöä.
Hyönteiset ovat hyvin monimuotoinen ja elinympäristövaatimuksiltaan vaihteleva eliölajiryhmä, joka tunnetaan osin huonosti. Hyönteisten suurta osuutta uhanalaisten suolajien määrästä selittää erityisesti ensisijaisesti suolla esiintyvien uhanalaisten perhosten (25 lajia) ja kaksisiipisten (16 lajia) suuri määrä (Hyvärinen ym. 2019).
Alueellisesti uhanalaisia lajeja on runsaasti erityisesti Suomen etelä- ja keskiosissa, kun taas Pohjois-Suomessa alueellisesti uhanalaisten lajien määrä on pienempi (kuvat 1 ja 2). Tärkein suolajien uhanalaisuuden syy ja tulevaisuuden uhkatekijä on luonnontilaisten suoelinympäristöjen väheneminen erityisesti metsäojituksen ja metsätaloustoimien takia (Hyvärinen ym. 2019).
Valtakunnallisesti uhanalaiset suoputkilokasvit
Valtakunnallisesti uhanalaisten suoputkilokasvien elinympäristöjä ovat erityisesti letot (taulukko 5). Suomessa lettojen esiintyminen painottuu kalkkipitoisen maaperän alueille. Tämä korostaa Ahvenanmaata ja Pohjois-Suomea valtakunnallisesti uhanalaisten suoputkilokasvien nykyesiintymien painopistealueina. Koska juuri maaperän kalkkipitoisuus on useille lettolajeille keskeinen vaatimus, niitä esiintyy myös muualla kuin soilla. Joitain lajeja esiintyy myös ihmisen luomissa kalkkipitoisissa elinympäristöissä. Lettojen lisäksi korvet ovat useiden uhanalaisten suoputkilokasvien elinympäristö. Ahvenanmaan ja muun lounaisen Suomen merkitystä korostaa kalkkipitoisen maaperän lisäksi se, että osa valtakunnallisesti uhanalaisista lajeista on levinneisyydeltään eteläisiä. Ne eivät tule toimeen ilmastoltaan kylmemmissä osissa maata.
Taulukko 5. Suoelinympäristöissä tavattavat valtakunnallisesti uhanalaiset (CR, EN, VU) putkilokasvit Suomessa (ensi- ja toissijaiset suolajit, Hyvärinen ym. 2019). Suoelinympäristöt jaotellaan keskustavaikutteisiin elinympäristöihin (letot, nevat, rämeet) ja reunavaikutteisiin elinympäristöihin (lähdesuot, luhdat, korvet). Häiriöympäristöt viittaavat tässä etupäässä ihmisen toiminnan tuloksena syntyneisiin avoimiin kasvupaikkoihin (ojanpenkat, tienvarret, jätemaakasat jne.). Alkuperäisen levinneisyyden laajuuden ääripäätapaukset on ilmaistu taulukossa merkeillä: █ melko laaja, joskin pohjoispainotteinen, vähintään noin 50 % Suomen alasta; ▄ Koko hemi- ja eteläboreaalinen alue; ▲ Ahvenanmaa tai pääosin Ahvenanmaa (ilmastollisesti ja kallioperältään Suomen hemiboreaalisen vyöhykkeen edullisinta osaa). Uhanalaisten kasvien luettelo on haettu Punaisen kirjan verkkopalvelusta (https://punainenkirja.laji.fi/) 17.2.2020. Uhanalaisuusluokat: CR = äärimmäisen uhanalaiset, EN = erittäin uhanalaiset, VU = vaarantuneet.
Alueellisesti uhanalaisten suoputkilokasvien tarkastelu (kuva 1) toisaalta korostaa voimakkaimmin ojitettua Etelä- ja Keski-Suomea Ahvenanmaan ja Pohjois-Suomen sijaan. Alueellisesti uhanalaiset putkilokasvit (Ympäristöministeriö 2015) ovat tyypillisesti laajalle levinneitä lajeja. Vaikka ne ovat taantuneet jollain alueella, niitä on säilynyt niin paljon toisilla alueilla, etteivät ne ole valtakunnallisesti uhanalaisia. Tämä näkyy suurena alueellisesti uhanalaisten lajien määränä voimakkaimmin ojitetuilla alueilla eli Etelä- ja Keski-Suomessa.
Seuraavassa on katsaus valtakunnallisesti uhanalaisten, suolla esiintyvien putkilokasvien kasvupaikkavaihteluun ja levinneisyyskuvaan (Ryttäri & Kettunen 1997, Ryttäri ym. 2012, Hyvärinen ym. 2019, Lampinen & Lahti 2019). Soiden kasvillisuutta on yleisemmin käsitelty Soiden kasvillisuus -tekstissä.
Keskustavaikutteisten kalkkilettojen (Kaakinen ym. 2018) lajiston alkuperäinen levinneisyyskuva (Ryttäri ym. 2012, Hyvärinen ym. 2019, Lampinen & Lahti 2019) Suomessa on kaksijakoinen (taulukko 5). Yhtäältä erottuu Kuusamoon ja Lapin kolmioon painottuva lajiryhmä (ryhmä 1). Ryhmän lajeista yhdellä (lapinkämmekkä) on laajempikin levinneisyysalue. Toisaalta Ahvenanmaalle ja Lapin kolmioon painottuu myös pienempi lajiryhmä (ryhmä 2), jonka lajeja on tavattu myös kalkkipitoisissa ihmisen luomissa häiriöympäristöissä. Kuusamolaisiin kalkkilettojen lajeihin voitaisiin yhdistää myös kuusamonsarake, joka on luokiteltu kalkkilähteikköjen (ei lainkaan soiden) lajeihin (Hyvärinen ym. 2019). Se kasvaa yhden ainoan lähdepuron reunalla paljaalla mineraalimaalla ja puronvarren ohutturpeisella letolla (Ryttäri ym. 2012).
Reunavaikutteisten, koivuletto- ja lähdelettotyyppisten lettojen (Kaakinen ym. 2018) lajien (ryhmät 3 ja 4) alkuperäisestä levinneisyyskuvasta (Ryttäri ym. 2012) kaksijakoisuus puuttuu (taulukko 5). Lajit edustavat suurelta osin laajaa, joskin pohjoispainotteista levinneisyyskuvaa. Pääosa lajeista (ryhmä 4) on samalla tyypillisesti lähteikköjen lajeja. Luhtalettojen edustus on yhden, pääosin eurooppalais-pohjoisamerikkalaisen lehtometsävyöhykkeen lajin (taarna) varassa (ryhmä 5). Reunavaikutteisten lettojen lajistoon kuuluu edelleen lettokorpien ja -rämeiden painanteiden himmeävilla (ryhmä 6), jota on tavattu myös häiriöympäristöissä. Sen alkuperäinen levinneisyys käsittää laajalti Pohjois-Suomen.
Lettojen osalta on edelleen huomionarvoista, että useimmat suolajit, jotka esiintyvät sekä letoilla että erilaisilla niityillä, esiintyvät myös häiriöympäristöissä (taulukko 5). Ne ovat lähinnä Ahvenanmaan lettojen vaateliaita saroja, esikkokasveja (jauhoesikko) ja kämmeköitä (ryhmät 7 ja 8). Peräpohjolassa ja Lapissa tavataan vastaavanlainen saramaisten kasvien uhanalaisryhmä (ryhmä 9) niittymäisillä joenrantasomerikoilla tai kosteilla tunturiniityillä ja letoilla. Niin ikään yksi kalkkivaikutteisten tunturikankaiden kämmekkä esiintyy myös letoilla (ryhmä 10).
Nevoillakin esiintyvät lajit (taulukko 5, ryhmä 11) edustavat kausivetisten, luontaisesti lievästi häirittyjen hemi- ja eteläboreaalisten nevaympäristöjen (Laitinen 2008) lajeja (pikkukihokki), ääri-ilmiönä mereinen kellokanerva ojituksen muuttamalla suolla.
Reunavaikutteiset, korpisuuden, luhtaisuuden ja lähteisyyden luonnehtimat suoelinympäristöt (Eurola ym. 2015) vaihettuvat suon ulkopuolisiin elinympäristöihin. Reunavaikutteisten suoelinympäristöjen lajien pääasialliset kasvupaikat ovatkin muualla kuin soilla, eli metsissä, rantojen tulvamailla ja pienvesien lähteiköissä (taulukko 5; Hyvärinen ym. 2019). Nämä lajit voidaan suoelinympäristönsä perusteella ryhmitellä toisaalta laajalle levinneisiin ja ahvenanmaalaisiin metsien ja rehevien korpimaisten notkelmien lajeihin (ryhmä 12) ja toisaalta varsin eteläisiin, erilaisten tulvarantojen ja tervalepikkojen (tervaleppäluhtien) (ryhmä 13) ja lähteikköjen ja rehevien korpipainanteiden (ryhmä 14) lajeihin.
Soiden lintulajisto ja sen uhanalaisuus
Ympäristötekijöiden vaikutus lintulajiston esiintymiseen soilla
Lintulajien esiintyminen suolla riippuu ennen muuta suon puustoisuudesta ja märkyydestä (Fraixedas ym. 2017, 2020). Myös suon maantieteellisellä sijainnilla sekä lajien välisillä suhteilla on merkitystä (Järvinen ym. 1987, Väisänen ym. 1998). Harva- ja matalapuustoisilla rämeillä menestyvät monet avosoiden lajit, tiheäpuustoisilla soilla metsälajit tulevat vallitseviksi. Tämän vuoksi ojituksen vaikutus rämeiden ja soiden reunaosien linnustoon näkyy vasta hyvin pitkällä, vuosien tai jopa vuosikymmenten viiveellä. Puustoisilla soilla myös lehtipuiden (hieskoivun) määrä vaikuttaa lintulajistoon. Märillä avosoilla linnusto on runsaampi kuin kuivilla. Avovesipintaisilla soilla esiintyy myös enemmän vesi- ja lokkilintuja kuin sammalpeitteisillä märillä soilla. Lisäksi mätäs-, väli- ja rimpipintojen vaihtelu runsastuttaa linnustoa.
Soiden linnusto on Suomen alueella keskimäärin runsaampi pohjoisessa kuin etelässä (Järvinen & Sammalisto 1976, Järvinen ym. 1987). Tämä poikkeaa maailmanlaajuisella tasolla havaitusta ja yleisesti odotetusta biologisesta säännönmukaisuudesta, jonka mukaan samaa tyyppiä edustavien elinympäristöjen lajimäärät vähenevät etelästä pohjoiseen, joskin alle 20 leveysasteen etäisyyksillä tunnetaan myös päinvastaisia trendejä (Willig ym. 2003). Suomen soiden epätavallinen trendi selittyy sillä, että kaikki suolajit riekkoa lukuun ottamatta ovat muuttolintuja, jotka käyvät ainoastaan hyödyntämässä lisääntymiskaudellaan kesän erittäin runsasta ravinnon tarjontaa Pohjois-Suomen märillä ja rimpisillä aapasoilla. Linnusto on myös muuttunut siten, että pohjoisilla soilla on yhä suurempi lajimäärä suhteessa eteläisiin soihin (Fraixedas ym. 2020). Muutoksen syynä on erityisesti se, että ojituksen ja alun perinkin vähäisemmän linnuille otollisen avosuopinta-alan johdosta eteläisessä Suomessa on soiden linnuille soveltuvia soita niin vähän ja kaukana toisistaan, etteivät ne enää pysty ylläpitämään elinvoimaisia populaatioita (Fraixedas ym. 2020). Petolintujen, erityisesti muuttohaukan, saalistus voi keski- ja pohjoisboreaalisella vyöhykkeellä paikallisesti vaikuttaa pesimäsuon linnustoon, etenkin kahlaajiin.
Soiden uhanalaiset lintulajit Suomessa ja ojituksen vaikutus suolinnustoon
Linnusto on eliöryhmä, joka tunnetaan Suomessa erittäin hyvin (Lehikoinen ym. 2019), jopa kaikkein parhaiten eri eliöryhmistä. Lintujen esiintymisestä ja määristä on paljon aineistoja pitkältä ajalta (esim. Väisänen & Rauhala 1983, Väisänen 2006, Fraixedas ym. 2017). Uusimmat arvioinnit lintulajien uhanalaisuudesta ovat vuosilta 2010 (Mikkola-Roos ym. 2010) ja 2019 (Lehikoinen ym. 2019) ja väliarvio vuodelta 2015 (Tiainen ym. 2016a).
Soiden ojituksen on havaittu vähentävän selvästi monien suolintulajien yksilömääriä; ainoana poikkeuksena on pohjansirkku, joka on reunaelinympäristöjen (korpien ja rämeiden) laji (Fraixedas ym. 2017). Sekin on arvioitu silmälläpidettäväksi (Lehikoinen ym. 2019). Ojitettujen korpien ja rämeiden muuttuessa vähitellen kangasmetsien kaltaiseksi pohjansirkku todennäköisesti häviää niiltä, joskin varvikon rehevöityessä se voi tilapäisesti hyötyä ojituksesta (Ukkonen 1983). Lehikoisen ym. (2019) mukaan useimmat ensisijaisesti soilla esiintyvistä uhanalaisista ja silmälläpidettävistä lintulajeista ovat avosoiden lajeja, suokukko niistä uhanalaisimpana (taulukko 6). Lähes kaikki ensisijaiset suolajit ovat etupäässä aapasuovyöhykkeen lajeja. Toissijaisissa suolajeissa on paljon myös eteläisessä Suomessa esiintyviä avoimien alueiden lajeja. Avosoiden ja rämeiden uhanalaisiksi luokiteltuihin lajeihin kuuluu taigametsähanhi, joka pesii rämeillä käyttäen avosoita sulkimiseen ja ravinnonhankintaan.
Taulukko 6. Ensisijaisesti ja toissijaisesti suolla esiintyvät uhanalaiset (CR, EN, VU) ja silmälläpidettävät (NT) lintulajit suoelinympäristöittäin uhanalaisuusluokan mukaisessa järjestyksessä (Hyvärinen ym. 2019). Tiedot on haettu Punaisen kirjan verkkopalvelusta (https://punainenkirja.laji.fi/) 17.2.2020. Taulukossa on lajin nimen perässä lajin levinneisyysalue Suomessa karkeasti suovyöhykkeiden avulla luonnehdittuna etelästä pohjoiseen: K Etelä-Suomen keidassuoalue, A Pohjois-Suomen aapasuoalue, P Pohjois-Lapin palsasuoalue. Uhanalaisuuteen vaikuttaneet, ojitukseen ja turpeenottoon liittyvät syyt: O/o ojitus ja turpeenotto ovat ensisijainen/yksi uhanalaisuuden syy, – ojitusta ei ole luokiteltu uhanalaisuuden syyksi, ? uhanalaisuuden syy tuntematon. Uhanalaisuusluokat: CR = äärimmäisen uhanalaiset, EN = erittäin uhanalaiset, VU -= vaarantuneet
Ojitusta ei ole arvioitu ensisijaiseksi uhanalaistumisen syyksi kaikkein uhanalaisimmilla avosoiden lintulajeilla (Lehikoinen ym. 2019). Muuttohaukan uhanalaistumiseen ovat johtaneet ensisijaisesti ympäristömyrkyt ja metsästys, ja muuttohaukkakanta onkin elpynyt suojelutyön ansiosta (Lehikoinen ym. 2019). Suokukon uhanalaistumiseen syynä ovat ennen kaikkea muutokset Suomen ulkopuolella, mutta myös kosteiden niittyjen ja muiden avointen kosteikkoalueiden kuin soiden umpeenkasvu Suomessa. Rämeiden ja soistuneiden kankaiden uhanalaiselle riekolle ojitus on kuitenkin arvioitu ensisijaiseksi uhanalaistumisen syyksi, joskin myös lumipeitteisen ajan lyheneminen aiheuttaa riekolle ongelmia (Melin ym. 2020). Sinänsä hyvin tunnetuissa linnuissa on suolajeissakin useita lajeja, kuten vesipääsky, jotka ovat uhanalaistuneet, mutta joiden uhanalaistumisen syitä ei tunneta (taulukko 6).
Useimmat soiden uhanalaiset ja silmälläpidettävät lintulajit ovat levinneisyydeltään pohjoisia. Vuoden 2015 uhanalaisarvioinnissa uhanalaisten lajien osuus arvioidusta lajistosta oli soilla pienempi mutta silmälläpidettävien lajien osuus suurempi kuin kaikkien elinympäristöjen arvioidussa lajistossa keskimäärin (Tiainen ym. 2016b). Soilla uhanalaistuminen johtuu muita elinympäristöjä useammin siitä, että vaikka lajin yksilömäärä voi olla suurikin, se on pienentynyt voimakkaasti viime vuosina tai sen arvioidaan pienenevän voimakkaasti lähivuosina. Muissa elinympäristöissä, karuja järviä lukuun ottamatta, uhanalaistuminen johtuu soita useammin siitä, että lajin yksilömäärä on jo hyvin pieni ja pienenee edelleen (Tiainen ym. 2019).
Soiden hyönteislajisto ja sen uhanalaisuus
Ympäristötekijöiden vaikutus hyönteislajiston esiintymiseen soilla
Soiden hyönteislajiston elinympäristövaatimukset vaihtelevat paljon lajista ja lajiryhmästä toiseen. Myös tietämys asiasta on epätasaisesti puutteellista eri ryhmissä. Kauri Mikkola esitti Suo-lehdessä yli 40 vuotta sitten (Mikkola 1976) karkean mallin suohyönteisten jakautumisesta erilaisiin elinympäristöihin. Hänen hahmottelunsa mukaan (1) toukkina tai myös aikuisina matalissa vesissä, kostealla paljaalla turpeella ja rahkasammaleella elävän lajiston koostumukseen vaikuttavat suoveden happamuus (pH), happipitoisuus ja humuspitoisuus. Toisaalta (2) toukkina kasvien maanpäällisiä osia syöville tai kukissa aikuisina käyville hyönteisille merkityksellistä on sopivien ravintokasvien esiintymisen lisäksi äärevä pienilmasto ja sen aiheuttama suon viileys ympäristöön verrattuna. Tällainen äärevä pienilmasto vallitsee kuivapintaisissa suon osissa, erityisesti rämeillä, koska kuiva turve johtaa huonosti lämpöä. Muita ryhmiä ovat (3) edellisten pedot ja (4) muut. Esimerkkeinä tarkastellaan lyhyesti sääskiä, perhosia, muurahaisia ja sudenkorentoja.
Soiden vaaksiaiset kuuluvat sääskiin ja Mikkolan (1976) ykkösryhmään. Niillä lajimäärän on havaittu kasvavan kohoavan pH:n, joka korreloi suon ravinteisuuden kanssa (Tahvanainen 2004), sekä kasvillisuuden mätäs-väli-rimpipintavaihtelun ja suotyyppivaihtelun lisääntymisen myötä. Karut suonosat ovat kuitenkin tietyille lajeille merkittäviä elinympäristöjä (Salmela 2004). Lähteikköjen, soitten ja virtavesiympäristöjen vaaksiaisyhteisöjen ja lajimäärien perusteella pohjavesivaikutteisilla eli lähteisillä paikoilla on paljon vaaksiaisia. Tällöin pohjavesivaikutukseen liittyvä vakaavetisyys (kosteuden säilyminen kuivinakin kesinä) saattaisi olla merkittävä seikka vaaksiaislajirunsaudelle (Salmela ym. 2007). Vaaksiaislajisto soilla riippuu siis melko pitkälle samantapaisista ympäristötekijöistä kuin kasvilajistokin.
Maantieteellinen sijainti vaikuttaa voimakkaasti vaaksiaislajistoon. Vaaksiaisilla yleisesti lajimäärä monien muiden hyönteisten tapaan (Väisänen & Heliövaara 1994) pienenee pohjoiseen päin (Salmela 2012), mikä pitää yhtä maailmanlaajuisella tasolla havaitun ja yleisesti odotetun säännönmukaisuuden (Willig ym. 2003) kanssa. Suovaaksiaisilla maantieteellinen ja ilmastollinen lajimäärän vaihtelu Suomessa on kuitenkin soiden pesimälintujen tapaan epätavallinen: lajeja on enemmän pohjoisessa kuin etelässä.
Soiden perhoset kuuluvat Mikkolan (1976) kakkosryhmään. Lähes kaikki suoperhoslajit ovat toukkavaiheessa moniruokaisia ja käyttävät ravintonaan useita yleisiä suokasveja (Seppänen 1970, Pöyry 2001). Ravintokasvien lisäksi suoperhosten esiintymistä rajoittavina tekijöinä ovat suon kosteus- ja lämpöolot (Mikkola 1976, Marttila 2005) sekä suota ympäröivien alueiden ominaisuudet, kuten metsäisyys tai avoimuus (Väisänen 1992). Suoperhoset ovat riippuvaisia omasta elinympäristöstään ja lentävät vain harvoin suon ulkopuolelle. Runsain perhoslajisto on kitukasvuista ja matalaa männikköä kasvavilla rämeillä. Tuulisilla avosoilla perhosia tavataan eniten suon reunoilla, kuten keidassoiden reunaluisilla ja laiteilla, vähiten suon keskellä (Väisänen 1992, Marttila ym. 2001, Noreika ym. 2016).
Suoympäristön uhanalaiset hyönteiset Suomessa ja ojituksen vaikutus soiden hyönteislajistoon
Soille erikoistuneet hyönteislajit ovat yleensä riippuvaisia kosteudesta ja avoimuudesta (Noreika ym. 2015). Avointen ja vähäpuustoisten karujen soiden metsäojitusta ja ennallistamista tutkittaessa havaittiin, että soille erikoistuneet hyönteiset (ja samoin hämähäkit) hyötyvät märistä kuljuista ja rimmistä, suurten puiden puuttumisesta ja rahkasammalista. Ojitus heikensi näitä tekijöitä ja samalla soille erikoistuneiden lajien elinolosuhteita. Ennallistaminen päinvastoin palautti osan lajeista jo muutamassa vuodessa. Soilla esiintyvien, monenlaisiin oloihin sopeutuneiden yleislajien esiintymiseen ojituksella tai ennallistamisella ei ollut selkeää vaikutusta (Noreika ym. 2015).
Suomessa esiintyy suuri määrä sääskilajeja, joihin kuuluvat esimerkiksi vaaksiaiset, surviaissääsket ja sienisääsket. Vuonna 2019 uhanalaisarvioita tehtäessä tunnettujen lajien määrä oli 2980, ja läheskään kaikkia lajeja ei vielä tunneta (Salmela ym. 2019). Arvioiduista lajeista 79 % on luokiteltu elinvoimaisiin (LC), 4 % uhanalaisiin (CR, EN, VU), 7 % silmälläpidettäviin (NT) ja 10 % puutteellisesti tunnettuihin (DD). Uhanalaisissa vaaksiaislajeissa on metsälajien lisäksi pienvesien ja soiden lajeja. Uhanalaisilla surviaissääskilajeilla toukkien elinympäristöinä ovat lähteikköjen lisäksi muun muassa letot ja palsasuot. Uhanalaiset kuivan maan sienisääskilajit sen sijaan ovat vanhojen metsien ja lehtojen ja tuoreimpien havaintojen mukaan (Salmela ym. 2015) myös lettojen lajeja. Ojituksen vaikutuksesta sääskilajistoon on Suomessa tehty yksi tutkimus vaaksiaisista (Noreika ym. 2015). Sen mukaan soihin erikoistuneet vaaksiaiset ja niiden yhteisöt kärsivät ojituksesta eli käytännöllisesti katsoen puuttuivat ojitetuilta soilta. Ojituksella vaikuttaisi siis olevan hyvin merkittävä vaikutus ainakin tämän sääskiryhmän soilla esiintyvään lajistoon (Salmela ym. 2019).
Perhoset on Suomessa perinteisesti paljon harrastettu ryhmä, jonka tuntemuksen taso on erinomainen (Nupponen ym. 2019). Myös suoperhosten uhanalaisuutta ja suojelutilannetta Etelä-Suomen soilla on arvioitu (Pöyry 2001). Suomesta tunnetaan 2600 perhoslajia (Nupponen ym. 2019). Näistä 58 lajia elää pelkästään soilla (Pöyry 2001). Metsäojituksesta johtuva elinympäristöjen pirstoutuminen ja heikentyminen on vaikuttanut etenkin levinneisyydeltään eteläisiin suoperhoslajeihin, koska soiden ojitus on ollut voimakasta Etelä-Suomen keidassuovyöhykkeellä (Pöyry 2001, Marttila 2005). Suoperhoset taantuvat ojituksen seurauksena (Noreika ym. 2016). Suoperhosten yksilö- ja lajimäärä oli ojitetuilla soilla pienempi kuin luonnontilaisilla. Syynä olivat sekä toukkien ravintokasvien vähyys että kookas puusto.
Suot ovat muurahaisille keskeinen elinympäristö. Suomessa esiintyvistä muurahaislajeista suot ovat kymmenesosalle ensisijainen ja kolmasosalle tärkeä elinympäristö (Punttila ym. 2013). Soiden merkitystä korostaa se, että erilaisilla soilla – korvissa, rämeillä, nevoilla ja letoilla – on erilainen muurahaisyhteisö. Esimerkiksi suolajeista suomustahainen (Formica picea), mustapäämuurahainen (Formica uralensis) ja rämeloviniska (Formica forsslundi) esiintyvät puustoisilla soilla lähes yksinomaan rämeillä (Punttila & Kilpeläinen 2009, Punttila ym. 2013). Avosuolta puuttuvat monien kekomuurahaisten ravintotaloudelle tärkeät puissa elävät kirvat ja maaperän vetisyys estää syvällä maan alla talvehtimisen, kun taas karulla rämeellä olosuhteet ovat monille muurahaislajeille jopa kivennäismaita otollisemmat (Vepsäläinen ym. 2000, Punttila & Kilpeläinen 2009). Rämeet korostuvatkin runsaslajisina muurahaisparatiiseina, kun taas korvet ja avosuot ovat selvästi vähälajisempia (Punttila ym. 2013).
Ojitus johtaa vähitellen soille erikoistuneiden muurahaislajien häviämiseen, vaikka osa lajeista saattaa aluksi jopa hyötyä ojituksesta (Vepsäläinen ym. 2000, Punttila & Kilpeläinen 2009, Punttila ym. 2016). Kivennäismailla ja puustoisilla soilla yleiset metsälajit valtaavat vähitellen metsäojitetut suot varjostuksen lisääntyessä ja suon kehittyessä kohti turvekangasta (Vepsäläinen ym. 2000, Punttila & Kilpeläinen 2009, Punttila ym. 2016). Yhtään ensisijaisesti soilla esiintyvää muurahaislajia ei ole arvioitu uhanalaiseksi (Hyvärinen ym. 2019), mutta arviointia hankaloittaa se, että soiden muurahaiset tunnetaan vielä huonosti (Punttila ym. 2013).
Suomesta on tavattu 62 sudenkorentolajia, joista 54 kuuluu vakituiseen lajistoon (Penttinen & Mäkinen 2013, Karjalainen 2019). Monet lajit ovat ilmastonmuutoksen myötä levinneet aiempaa pohjoisemmaksi. Koska sudenkorentojen toukat elävät vedessä, soiden rimmet, allikot ja lammet ovat monille lajeille merkittäviä elinympäristöjä (Karjalainen 2002). Aikuisille sudenkorennoille avoimet ja aurinkoiset suot ovat myös otollisia saalistusalueita (Penttinen & Mäkinen 2013). Ojitetut suot eivät ole sudenkorennoille kovin otollisia elinympäristöjä, koska niillä on niukasti avovesipintoja ja avoimia elinympäristöjä (Elo ym. 2015). Joidenkin suolajien onkin havaittu taantuneen Etelä- ja Keski-Suomessa viime vuosikymmeninä (Penttinen & Mäkinen 2013). Soiden sudenkorennot esiintyvät kuitenkin yleensä myös matalissa vesistöissä, eikä ojitus ole johtanut suolla esiintyvien lajien uhanalaistumiseen (Karjalainen 2019). Jos soiden ennallistaminen palauttaa rimpiä ja allikoita, sudenkorennot voivat palata lisääntymään niihin hyvinkin nopeasti, koska aikuiset sudenkorennot lentävät pitkiäkin matkoja (Penttinen & Mäkinen 2013, Elo ym. 2015).
Alueellisesti uhanalaiset suolajit
Alueellinen uhanalaisuus voidaan arvioida lajeille, jotka on valtakunnallisessa uhanalaisuusarvioinnissa luokitettu silmälläpidettäviksi (NT) tai elinvoimaisiksi (LC). Alueellisesti uhanalaisten suolajien tarkastelu täydentää valtakunnallisesti uhanalaisten lajien antamaa kuvaa suolajiston uhanalaisuudesta Suomessa. Viimeisimmät tulokset lajien valtakunnallisesta uhanalaisuudesta julkaistiin vuonna 2019 (Hyvärinen ym. 2019). Lajien alueellisen uhanalaisuuden arviointi tullaan uusimaan valtakunnallisen arvioinnin valmistumisen jälkeen osalle lajiryhmiä, mutta nämä tulokset eivät ole vielä tätä kirjoitettaessa valmistuneet.
Tässä esitetyt tulokset alueellisesti uhanalaisten suoputkilokasvien ja -sammalten määristä perustuvat aikaisempiin arviointeihin (valtakunnallinen arviointi Rassi ym. 2010, alueelliset arvioinnit putkilokasveille Ryttäri ym. 2012 ja sammalille Sammaltyöryhmä 2017). Alueellisesti uhanalaisten suolajien lista noudattelee pääpiirteessään julkaisussa Ympäristöministeriö (2015; Liite V) esitettyä lajilistaa sillä poikkeuksella, että sammalten osalta on tässä yhteydessä käytetty päivitettyjä tuloksia vuodelta 2017.
Alueellisesti uhanalaisia suoputkilokasvilajeja on yhteensä 121, joista 50 on ensisijaisia suolajeja ja 71 toissijaisia suolajeja. Alueellisesti uhanalaisia suosammallajeja on yhteensä 88, joista 48 on ensisijaisia ja 40 toissijaisia suolajeja.
Alueellisesti uhanalaisten putkilokasvien (kuva 1a) ja sammalten (kuva 1b) määrä on suurempi maan etelä- ja keskiosissa kuin pohjoisosassa. Tähän vaikuttanee soiden voimakkaampi käyttö, kuten ojitus, hemi-, etelä- ja keskiboreaalisella vyöhykkeellä verrattuna pohjoisboreaaliseen vyöhykkeeseen. Alueellisesti uhanalaisten suosammalten määrä (kuva 1b) antaa Suomesta vielä putkilokasveja selvemmin kahtiajakoisen kuvan, joka on melko samanlainen kuin suoluontotyyppien antama kuva suoluonnon uhanalaisuudesta (ks. Suoluontotyyppien uhanalaisuus; Kaakinen ym. 2018). Kaikkein suurin alueellisesti uhanalaisten lajien määrä on keskiboreaalisella Pohjanmaalla (3a). Kyseessä on Suomen suovaltaisin alue (ks. Soiden käyttö Suomessa, kuva 3), jota leimaavat karut, tasaiset ja suuret suot, keidassuot ja Pohjanmaan aapasuot. Samalla boreaalinen Pohjanmaa kuuluu tehokkaimmin ojitettuun Suomen osaan. Paitsi erot maankäytön voimakkuudessa, alueellisesti uhanalaisten lajien määrän alueellista tilannetta voi osaltaan selittää myös Suomen luonnonvaraisten putkilokasvien lajimäärän väheneminen pohjoiseen päin (Suominen & Hämet-Ahti 1987, Lahti ym. 1988). Myös metsäisten kasvillisuusvyöhykkeiden sammallajisto niukkenee Suomessa jonkin verran pohjoista kohden (Ulvinen ym. 2002), mutta alueellisesti uhanalaisissa suosammalissa eteläiset lajit eivät kuitenkaan erityisesti korostu (Ympäristöministeriö 2015). Syitä lajien uhanalaisuuden alueellisiin eroihin olisi hyvä tutkia tarkemmin varmempien johtopäätösten tekemiseksi.
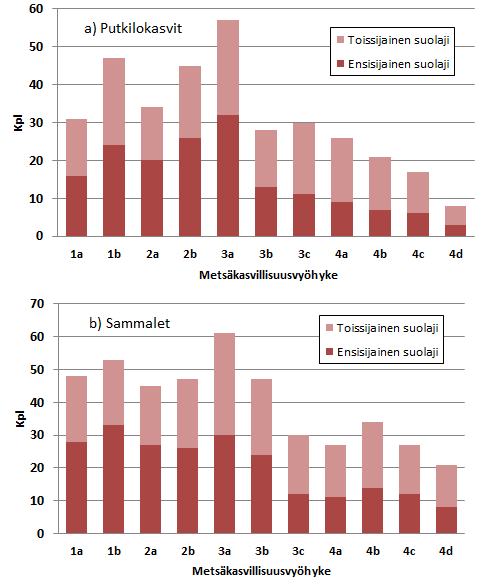
Kuva 1. a) Alueellisesti uhanalaisten suoputkilokasvien lukumäärä metsäkasvillisuusvyöhykkeiden lohkoilla. Alueelliset uhanalaisuusarviot Ryttärin ym. (2012) mukaan. b) Alueellisesti uhanalaisten sammalten lukumäärä metsäkasvillisuusvyöhykkeiden lohkoilla. Alueelliset uhanalaisuusarviot Sammaltyöryhmän (2017) mukaan. Kartta: Hyvärinen ym. 2019.
Linnuista tehtiin vuoden 2010 arvioinnin yhteydessä myös alueellinen uhanalaisuustarkastelu vuosille 2000 ja 2010 (Rajasärkkä ym. 2013). Valtakunnallisesti elinvoimaisiksi tai silmälläpidettäviksi vuonna 2000 luokitelluista suolajeista metsähanhi, sinisuohaukka, riekko, kurki, jänkäsirriäinen, suokukko, taivaanvuohi, pikkukuovi, liro, keltavästäräkki ja isolepinkäinen luokiteltiin osassa maata, yleensä eteläisillä metsäkasvillisuusvyöhykkeillä, alueellisesti uhanalaisiksi. Vuonna 2010 alueellisesti uhanalaisten listalta oli poistettu kurki ja taivaanvuohi (elinvoimaisia koko maassa) sekä sinisuohaukka, suokukko ja keltavästäräkki, jotka arvioitiin tuolloin valtakunnallisesti uhanalaisiksi. Alueellisesti uhanalaisten listalla oli uutena kapustarinta. Vuonna 2019 ei alueellista tarkastelua tehty erikseen, mutta metsähanhi ja riekko luokiteltiin valtakunnallisesti uhanalaisiksi ja siten ne poistuivat alueelliselta listalta (Lehikoinen ym. 2019, Tiainen ym. 2019). Alueellisesti uhanalaisia ovat siten tällä hetkellä jänkäsirriäinen, pikkukuovi, liro, isolepinkäinen ja kapustarinta.
Kiitokset
Kiitämme Olli Autiota (Etelä-Pohjanmaan elinkeino-, liikenne- ja ympäristökeskus), Olli Loukolaa (Oulun yliopisto), Ari Rajasärkkää (Metsähallitus, Luontopalvelut, Oulu), Jukka Salmelaa (Lapin maakuntamuseo) ja Tauno Ulvista (Oulun yliopisto) arvokkaista tiedoista ja kommenteista.
Kommentoineet
Sakari Sarkkola
Kirjallisuus
Aapala, K. 2001a. Korpien ekologiset ominaispiirteet ja suojelutilanne. Suomen ympäristö 490: 87–148. Saatavissa: http://hdl.handle.net/10138/40594
Aapala, K. 2001b. Soiden uhanalainen lajisto. Suomen ympäristö 490: 149–182. Saatavissa: http://hdl.handle.net/10138/40594
Elo, M., Penttinen, J. & Kotiaho, J.S. 2015. The effect of peatland drainage and restoration on Odonata species richness and abundance. BMC Ecology 15: Article 11. https://doi.org/10.1186/s12898-015-0042-z
Eurola, S., Huttunen, A., Kaakinen, E., Kukko-oja, K., Saari, V. & Salonen, V. 2015. Sata suotyyppiä. Opas Suomen suokasvillisuuden tuntemiseen. Thule-instituutti, Oulangan tutkimusasema, Oulun yliopisto. 112 s. ISBN 978-952-62-0891-6
Fraixedas, S., Lindén, A., Husby, M. & Lehikoinen, A. 2020. Declining peatland bird numbers are not consistent with the increasing Common Crane population. Journal of Ornithology 161: 691–700. Saatavissa: https://doi.org/10.1007/s10336-020-01777-6
Fraixedas, S., Lindén, A., Meller, K., Lindström, Å., Keišs, O., Kålås, J.A., Husby, M., Leivits, A., Leivits, M. & Lehikoinen, A. 2017. Substantial decline of Northern European peatland bird populations: Consequences of drainage. Biological Conservation 214: 223–232. Saatavissa: https://doi.org/10.1016/j.biocon.2017.08.025
Grime, J.P. 1977. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. The American Naturalist 111: 1169–1194. Saatavissa: https://www.jstor.org/stable/2460262
Grime, J.P. 1979. Plant strategies and vegetation process. John Wiley & Sons, Chichester. 222 s.
Hyvärinen, E., Juslén, A., Kemppainen, E., Uddström, A. & Liukko, U.-M. (toim.) 2019. Suomen lajien uhanalaisuus – Punainen kirja 2019. Ympäristöministeriö & Suomen ympäristökeskus. Helsinki. 704 s. Saatavissa: http://hdl.handle.net/10138/299501
Ilvessalo, Y. 1957. Suomen suot. Valtakunnan metsien inventointiin perustuva kuvaus. Suo 8(5): 51–61. Saatavissa: http://www.suo.fi/article/9222
Järvinen, O., Kouki, J. & Häyrinen, U. 1987. Reversed latitudinal gradients in total density and species richness of birds breeding on Finnish mires. Ornis Fennica 64: 67–73. Saatavissa: https://lintulehti.birdlife.fi:8443/pdf/artikkelit/658/tiedosto/of_64_67-73_artikkelit_658.pdf
Järvinen, O. & Sammalisto, L. 1976. Regional trends in the avifauna of Finnish peatland bogs. Annales Zoologici Fennici 13: 31–43. Saatavissa: https://www.jstor.org/stable/23731782
Kaakinen, E., Kokko, A., Aapala, K., Autio, O., Eurola, S., Hotanen, J.-P., Kondelin, H., Lindholm, T., Nousiainen, H., Rehell, S., Ruuhijärvi, R., Sallantaus, T., Salminen, P., Tahvanainen, T., Tuominen, S., Turunen, J., Vasander, H. & Virtanen, K. 2018. Suot. Teoksessa: Kontula, T. & Raunio, A. (toim.). Suomen luontotyyppien uhanalaisuus 2018. Osa I – tulokset ja arvioinnin perusteet. Suomen ympäristö 5/ 2018. s. 117–170. Saatavissa: http://urn.fi/URN:ISBN:978-952-11-4816-3
Karjalainen, S. 2019. Sudenkorennot – Dragonfiles and Damselflies, Odonata. Teoksessa: Hyvärinen, E., Juslén, A., Kemppainen, E. Uddström, A. & Liukko, U.-M. 2019 (toim.). Suomen lajien uhanalaisuus – Punainen kirja 2019. Ympäristöministeriö & Suomen ympäristökeskus. Helsinki. s. 355–357. Saatavissa: http://hdl.handle.net/10138/299501
Karjalainen, S. 2002. Suomen sudenkorennot. Tammi, Helsinki. 222 s.
Korhonen, K.T., Ihalainen, A. Ahola, A., Heikkinen, J., Henttonen, H.M., Hotanen, J.-P., Nevalainen, S., Pitkänen, J., Strandström, M. & Viiri, H. Suomen metsät 2009–2013 ja niiden kehitys 1921–2013. Luonnonvara- ja biotalouden tutkimus 59/2017. 86 s. Saatavissa: http://urn.fi/URN:ISBN:978-952-326-467-0
Lahti, T., Kurtto, A. & Väisänen, R.A. 1988. Floristic composition and regional species richness of vascular plants in Finland. Annales Botanici Fennici 25: 281–291. Saatavissa: https://www.jstor.org/stable/23725506
Laitinen, J. 2008. Vegetational and landscape level responses to water level fluctuations in Finnish, mid-boreal aapa mire–aro wetland environments. Universitatis Ouluensis A 513. Saatavissa: http://urn.fi/urn:isbn:9789514288791
Lampinen, R. & Lahti, T. 2019. Kasviatlas 2018. Helsingin yliopisto, Luonnontieteellinen keskusmuseo, Helsinki. Levinneisyyskartat osoitteessa http://koivu.luomus.fi
Lehikoinen, A., Jukarainen, A., Mikkola-Roos, M., Below, A., Lehtiniemi, T., Pessa, J., Rajasärkkä, A., Rintala, J., Rusanen, P., Sirkiä, P., Tiainen, J. & Valkama, J. 2019. Linnut. Teoksessa: Hyvärinen, E., Juslén, A., Kemppainen, E., Uddström, A. & Liukko, U.-M. (toim.). Suomen lajien uhanalaisuus – Punainen kirja 2019. Ympäristöministeriö & Suomen ympäristökeskus, Helsinki. s. 560–570. Saatavissa: http://hdl.handle.net/10138/299501
Marttila, O. 2005. Suomen päiväperhoset elinympäristössään. Käsikirja. Auris, Joutseno. 272 s.
Marttila, O., Saarinen, K., Aarnio, H., Haahtela, T. & Ojalainen, P. 2001. Päiväperhosopas. Suomi ja lähialueet. Tammi, Helsinki. 231 s.
Melin, M., Mehtätalo, L., Helle, P., Ikonen, K. & Packalen, T. 2020. Decline of the boreal willow grouse (Lagopus lagopus) has been accelerated by more frequent snow-free springs. Scientific Reports 10: 6987 Saatavissa: https://doi.org/10.1038/s41598-020-63993-7.
Mikkola, K. 1976. Piirteitä soiden hyönteisten ekologiasta. Suo 27(1): 3–8. Saatavissa: http://www.suo.fi/article/9450
Mikkola-Roos, M., Tiainen, J., Below, A., Hario, M., Lehikoinen, A., Lehikoinen, E., Lehtiniemi, T., Rajasärkkä, A., Valkama, J. & Väisänen, R. A. 2010. Linnut. Teoksessa: Rassi, P., Hyvärinen, E., Juslén, A. & Mannerkoski, I. (toim.). Suomen lajien uhanalaisuus – Punainen kirja 2010. Ympäristöministeriö & Suomen ympäristökeskus, Helsinki. s. 320–331. Saatavissa: http://hdl.handle.net/10138/299499
Noreika, N., Kotiaho, J., Penttinen, J., Punttila, P., Vuori, A. Pajunen, T., Autio, O., Loukola, O. & Kotze, J. 2015. Rapid recovery of invertebrate communities after ecological restoration of boreal mires. Restoration Ecology 23(5): 566–579. Saatavissa: https://doi.org/10.1111/rec.12237
Noreika, N., Kotze, D.J., Loukola, O.J., Sormunen, N., Vuori, A., Päivinen, J., Penttinen, J., Punttila, P. & Kotiaho, J.S. 2016. Specialist butterflies benefit most from the ecological restoration of mires. Biological Conservation 196: 103–114. Saatavissa: https://doi.org/10.1016/j.biocon.2016.02.014
Nupponen, K., Nieminen, M., Kaitila, J.-P., Hirvonen P., Leinonen R., Koski H., Kullberg J., Laasonen E., Pöyry J., Sallien T. & Välimäki P. 2019. Perhoset. Teoksessa: Hyvärinen E., Juslén A., Kemppainen E. Uddström A. & Liukko U.-M. (toim.). Suomen lajien uhanalaisuus, Punainen kirja 2019. Ympäristöministeriö & Suomen ympäristökeskus. Helsinki. s. 470–508. Saatavissa: http://hdl.handle.net/10138/299501
Penttinen, J. & Mäkinen, J. 2013. Soiden sudenkorennot. Teoksessa: Aapala K., Similä M., Penttinen J. (toim.). Ojitettujen soiden ennallistamisopas. Metsähallituksen luonnonsuojelujulkaisuja. Sarja B 188. s. 93–94. Saatavissa: https://julkaisut.metsa.fi/julkaisut/show/1601
Punttila, P. & Kilpeläinen, J. 2009. Distribution of mound-building ant species (Formica spp., Hymenoptera) in Finland: preliminary results of a national survey. Annales Zoologici Fennici 46: 1–15. Saatavissa: http://www.sekj.org/PDF/anz46-free/anz46-001.pdf
Punttila, P., Vepsäläinen, K. & Väänänen, S. 2013. Muurahaiset, soiden ojitus ja ennallistaminen. Teoksessa: Aapala K., Similä M., Penttinen J. (toim.). Ojitettujen soiden ennallistamisopas. Metsähallituksen luonnonsuojelujulkaisuja. Sarja B 188. s. 86–90. Saatavissa: https://julkaisut.metsa.fi/julkaisut/show/1601
Punttila, P., Autio, O., Kotiaho, J.S., Kotze, D.J., Loukola, O.J., Noreika, N., Vuori, A. & Vepsäläinen, K. 2016. The effects of drainage and restoration of pine mires on habitat structure, vegetation and ants. Silva Fennica 50(2); article 1462. Saatavissa: https://doi.org/10.14214/sf.1462
Pöyry, J. 2001. Suoperhosten uhanalaisuus ja suojelutilanne Etelä-Suomessa. Teoksessa: Aapala, K. (toim.). Soidensuojelualueverkon arviointi. Suomen ympäristö 490. s. 213–257. Saatavissa: http://hdl.handle.net/10138/40594
Rajasärkkä, A., Below, A., Hario, M., Lehikoinen, A., Lehikoinen, E., Lehtiniemi, T., Mikkola-Roos, M., Tiainen, J., Valkama, J. & Väisänen, R. A. 2013. Lintujen alueellinen uhanalaisuus Suomessa. Linnut-vuosikirja 2012: 44–49. Saatavissa: https://lintulehti.birdlife.fi:8443/pdf/artikkelit/1831/tiedosto/Linnut_VK2012_044-049_Alue-UHEX_artikkelit_1831.pdf
Rassi, P., Hyvärinen, E., Juslén, A. & Mannerkoski, I. (toim.) 2010. Suomen lajien uhanalaisuus – Punainen kirja 2010. Ympäristöministeriö & Suomen ympäristökeskus, Helsinki. 685 s. Saatavissa: http://hdl.handle.net/10138/299499
Ryttäri, T., Kalliovirta, M. & Lampinen R. (toim.) 2012. Suomen uhanalaiset kasvit. Tammi, Helsinki. 384 s.
Ryttäri, T. & Kettunen, T. 1997. Uhanalaiset kasvimme. Kirjayhtymä, Helsinki. 335 s.
Salmela, J. 2004. Semiaquatic flies (Diptera, Nematocera) of three mires in the southern boreal zone, Finland. Memoranda Societatis pro Fauna et Flora Fennica 80: 1–10.
Salmela, J. 2012. Biogeographic Patterns of Finnish Crane Flies (Diptera, Tipuloidea). Psyche: A Journal of Entomology 2012: Article 913710: 1–20. Saatavissa: https://doi.org/10.1155/2012/913710
Salmela, J., Autio, O. & Ilmonen, J. 2007. A survey on the nematoceran (Diptera) communities of southern Finnish wetlands. Memoranda Societatis pro Fauna et Flora Fennica 83: 33–47.
Salmela, J., Siivonen, S., Dominiak, P., Haarto, P., Heller, K., Kanervo, J., Martikainen, P., Mäkilä, M., Paasivirta, L., Rinne, A., Salokannel, J., Söderman, G. & Vilkamaa, P. 2015. Malaise-hyönteispyynti Lapin suojelualueilla 2012–2014. Metsähallituksen luonnonsuojelujulkaisuja. Sarja A 221. 141 s. Saatavissa: https://julkaisut.metsa.fi/assets/pdf/lp/Asarja/a221.pdf
Salmela, J., Jakovlev, J., Paasivirta, L., Ilmonen, J., Kakko, I. & Haarto, A. 2019. Sääsket. Teoksessa: Hyvärinen E., Juslén A., Kemppainen E., Uddström A. & Liukko U.-M. (toim.). Suomen lajien uhanalaisuus – Punainen kirja 2019. s. 514–531. Saatavissa: http://hdl.handle.net/10138/299501
Sammaltyöryhmä 2017: Suomen sammalien levinneisyys metsäkasvillisuusvyöhykkeissä ja ELY-keskuksissa. Suomen ympäristökeskus. 3.1.2017. Saatavilla: http://www.ymparisto.fi/fiFI/Luonto/Lajit/Lajiensuojelutyo/Eliotyoryhmat/Sammaltyoryhma/Suomen_sammalet
Seppänen, E.J. 1970. Suurperhostoukkien ravintokasvit. Animalia Fennica 14: 1–179.
Suominen, J. & Hämet-Ahti, L. 1987. Väheneekö putkilokasvien määrä etelästä pohjoiseen? Lutukka 3: 17–20.
Tahvanainen, T. 2004. Water chemistry of mires in relation to the poor-rich vegetation gradient and contrasting geochemical zones of northeastern Fennoscandian Shield. Folia Geobotanica 39: 353–369. Saatavissa: https://doi.org/10.1007/BF02803208
Tiainen, J., Lehtiniemi, T., Lehikoinen, A., Jukarainen, A., Mikkola-Roos, M., Below, A., Pessa, J., Rajasärkkä, A., Rintala, J., Rusanen, P., Sirkiä, P. & Valkama, J. 2019. Suomen lintujen uhanalaisuus 2019. Linnut-vuosikirja 2018: 14–25.
Tiainen, J., Mikkola-Roos, M., Below, A., Jukarainen, A., Lehikoinen, A., Lehtiniemi, T., Pessa, J., Rajasärkkä, A., Rintala, J., Sirkiä, P. & Valkama, J. 2016a. Suomen lintujen uhanalaisuus 2015. Ympäristöministeriö & Suomen ympäristökeskus. 51 s. Saatavissa: http://hdl.handle.net/10138/159435
Tiainen, J., Mikkola-Roos, M., Below, A., Jukarainen, A., Lehikoinen, A., Lehtiniemi, T., Pessa, J., Rajasärkkä, A., Rintala, J., Sirkiä, P. & Valkama, J. 2016b. Suomen lintujen uhanalaisuus elinympäristöittäin. Linnut-vuosikirja 2015: 4–11.
Ukkonen, M. 1983. Pohjansirkun (Emberiza rustica) pesimäbiologiasta. Pro gradu. Helsingin yliopisto, matemaattis-luonnontieteellinen osasto, eläintieteen laitos. 210. s. Abstrakti saatavissa: http://hdl.handle.net/10138/32172
Ulvinen, T., Syrjänen, K. & Anttila, S. (toim.) 2002. Suomen sammalet – levinneisyys, ekologia, uhanalaisuus. Suomen ympäristö 560: 1–354. Saatavissa: http://hdl.handle.net/10138/40617
Vepsäläinen, K., Savolainen, R., Tiainen, J. & Vilén, J. 2000. Successional changes of ant assemblages: from virgin and ditched bogs to forests. Annales Zoologici Fennici 37: 135–149. Saatavissa: http://www.sekj.org/PDF/anzf37/anzf37-135p.pdf
Väisänen, R. 1992. Distribution and abundance of diurnal Lepidoptera on a raised bog in southern Finland. Annales Zoologi Fennici 29: 75–92. Saatavissa: http://www.sekj.org/PDF/anzf29/anz29-075-092.pdf
Väisänen, R.A. 2006. Maalinnuston kannanvaihtelut Etelä- ja Pohjois-Suomessa 1983–2005. Linnut-vuosikirja 2005: 83–98.
Väisänen, R. & Heliövaara, K. 1994. Hot-spots of insect diversity in northern Europe. Annales Zoologi Fennici 31: 71–81. Saatavissa: http://www.sekj.org/PDF/anzf31/anz31-071-081.pdf
Väisänen, R.A. & Rauhala, P. 1983. Succession of land bird communities on large areas of peatland drained for forestry. Annales Zoologi Fennici 20: 115–127. Saatavissa: https://www.jstor.org/stable/43923434
Väisänen, R.A., Lammi, E. & Koskimies, P. 1998. Muuttuva pesimälinnusto. Otava, Helsinki. 567 s.
Willig, M.R., Kaufman, D.M. & Stevens, R.D. 2003. Latitudinal gradients of biodiversity: pattern, process, scale, and synthesis. Annual Review of Ecology, Evolution and Systematics 34: 273–309. Saatavissa: https://doi.org/10.1146/annurev.ecolsys.34.012103.144032
Ympäristöministeriö 2015. Suot ja turvemaat maakuntakaavoituksessa. Ympäristöministeriö, Helsinki. Suomen ympäristö 7/2015. 112 s. Saatavilla: http://hdl.handle.net/10138/156305